Modèles de souris SLA et neurones moteurs spinaux
Aperçu de l'implication des motoneurones spinaux dans la progression de la maladie dans les modèles murins de sclérose latérale amyotrophique (SLA).
Cette ressource décrit:
- Les modèles murins de la SLA reproduisent-ils la pathologie du motoneurone spinal observée dans la maladie humaine?
- Quelles mesures peuvent être utilisées pour évaluer l'implication des motoneurones spinaux?
- Comment évaluer l'atteinte des motoneurones spinaux dans les modèles de SLA sur des coupes de tissus?
Les modèles murins de la SLA reproduisent-ils la pathologie du motoneurone spinal observée chez l'homme?
Une caractéristique essentielle d'un modèle animal utile de la SLA est qu'il reproduit plusieurs aspects de la maladie humaine qui peuvent être facilement mesurés (voir Modèles de souris SLA pour le développement de médicaments). La perte des motoneurones et la dégénérescence axonale, dendritique et synaptique sont les caractéristiques de la SLA (Salzinger, 2024). La dégénérescence des motoneurones entraîne une faiblesse et une atrophie des muscles squelettiques, et finalement la mort, généralement due à une insuffisance respiratoire (Hardiman, 2011). La SLA familiale représente environ 10 % des cas, les mutations les plus courantes se produisant dans les gènes C9ORF72, SOD1, FUS et TARDBP (Hardiman, 2017). Alors que les mutations observées dans le gène TARDBP sont rares (<1%), la protéine (TDP-43) codée par le gène TARDBP forme des agrégats cytoplasmiques dans environ 97% des cas de SLA (Shatunov, 2021). C'est pourquoi des modèles animaux de la SLA ont été développés par génie génétique à partir de ces gènes et d'autres.
Le modèle de souris transgénique hTDP-43ΔNLS (rNLS8) est un exemple illustratif d'un modèle de souris SLA qui récapitule les caractéristiques neuromotrices de la maladie humaine (Walker, 2015) [voir Souris TDP-43 ΔNLS (rNLS8) pour le développement de médicaments contre la SLA]. Ces souris surexpriment le TDP-43 humain avec un signal de localisation nucléaire (NLS) défectueux, ce qui entraîne une mauvaise localisation (avec des niveaux réduits d'expression nucléaire du TDP-43) et la formation d'agrégats cytoplasmiques. En ce qui concerne les motoneurones spinaux, ce modèle de souris a montré une perte de motoneurones rapides-fatigables (FF) avec une épargne relative des motoneurones lents, ainsi que des preuves de dénervation et de réinnervation de la jonction neuromusculaire (NMJ), comme observé dans la SLA humaine (Spiller, 2016; Hur, 2022).
Quelles mesures en cours de vie peuvent être utilisées pour évaluer l'implication des motoneurones spinaux?
La possibilité d'évaluer les effets de la progression de la maladie sur les motoneurones spinaux pendant la phase in-life d'une étude préclinique d'efficacité thérapeutique présente des avantages significatifs. Plusieurs approches sont disponibles, notamment la visualisation directe de la moelle épinière et les mesures quantitatives d'imagerie associées, ainsi que les mesures en aval résultant de la dénervation des muscles cibles.
Imagerie de la moelle épinière in vivo
L'imagerie non invasive, telle que l'imagerie par résonance magnétique (IRM) in vivo, peut permettre des mesures quantitatives qui reflètent l'état des motoneurones spinaux. Grolez et al. (Grolez, 2018) ont montré une diminution progressive de l'épaisseur de la moelle épinière cervicale chez les souris transgéniques SOD186R. Marcuzzo et al. (Marcuzzo, 2017) ont constaté des changements dépendants du temps dans la matière blanche et la matière grise de la moelle épinière via l'imagerie du tenseur de diffusion (DTI) longitudinale in vivo chez les souris G93A-SOD1. Figini et al. (Figini, 2016) ont optimisé les paramètres DTI pour permettre une détection robuste des différences entre les moelles épinières des souris G93A-SOD1 et de type sauvage. Gatto et ses collègues (Gatto, 2018) ont réalisé une DTI in vivo chez des souris YFP,G93A-SOD1, et ont constaté des changements significatifs dans l'anisotropie fractionnelle (FA), la diffusivité axiale (AD) et la diffusivité radiale (RD) chez les animaux du jour postnatal 80 (P80). Ces différences d'imagerie in vivo entre les souris G93A-SOD1 et les souris témoins ont été corrélées avec des changements dans l'intensité de la fluorescence axonale et les marqueurs cellulaires membranaires. Underwood et al. (Underwood, 2011) ont découvert que l'imagerie du tenseur de diffusion (DTI) pouvait détecter les lésions axonales de la moelle épinière in vivo, où les valeurs FA étaient réduites exclusivement dans les voies ventrales de la substance blanche de la moelle épinière lombaire des souris G93A-SOD1 par rapport aux compagnons de lit de type sauvage, cet effet devenant plus prononcé avec la progression de la maladie et en corrélation avec la gravité de la maladie. Les valeurs réduites de l'AF se limitaient aux voies de la substance blanche issues des neurones moteurs, alors que les fibres de la substance blanche sensorielle étaient préservées. En outre, des changements significatifs de la diffusivité axiale et radiale ont été associés à la dégénérescence axonale mesurée par microscopie électronique (ME).
Imagerie non invasive de l'atrophie musculaire
Au lieu d'évaluer les propriétés de diffusion de la moelle épinière elle-même, ce qui peut être difficile dans les modèles murins, la mesure de l'atrophie musculaire résultant de la dénervation due à la dégénérescence axonale/perte de motoneurones spinaux par imagerie non invasive est moins difficile et peut être traduite cliniquement. D'un point de vue clinique, Wilcox et al. (Wilcox, 2021) ont montré que l'IRM volumétrique démontre une reproductibilité, une réactivité temporelle et une corrélation avec les évaluations cliniques de la fonction musculaire, et ont suggéré que l'IRM volumétrique pourrait être une mesure de résultat potentielle de la réinnervation musculaire. Jenkins et ses collègues (Jenkins, 2013; Jenkins, 2018) ont utilisé l'IRM pour mesurer les changements volumétriques musculaires et d'autres paramètres d'imagerie (par exemple, le signal T2) dans la SLA. Dans une étude parcellaire, ce groupe a constaté une perte progressive du volume musculaire dans des muscles spécifiques chez des personnes atteintes de SLA chez qui la faiblesse clinique progressait (Jenkins, 2013). Klickovic et al. (Klickovic, 2024) ont rapporté un déclin significatif du volume de la main dominante et des muscles de la tête et du cou, ainsi que de la surface de section transversale (CSA) des membres inférieurs, au cours de la période d'observation maximale individuelle.
Les mesures longitudinales in vivo de l'atrophie musculaire sont possibles dans les modèles animaux en utilisant des modalités telles que l'IRM et la tomodensitométrie (CT). Marcuzzo et ses collègues (Marcuzzo, 2011) ont réalisé une analyse quantitative longitudinale par IRM chez des souris G93A-SOD1 et ont mis en évidence des pertes significatives du volume musculaire des membres postérieurs à partir de l'âge de 8 semaines. Pasetto et al. (Pasetto, 2018) et van der Heyden et al. (van der Heyden, 2020) ont rapporté la faisabilité de mesures micro-CT des membres postérieurs inférieurs chez la souris comme alternative à l'IRM.
Notre équipe à Biospective a tiré parti de l'imagerie micro-CT préclinique pour évaluer l'atrophie musculaire dans les modèles TDP-43 de la SLA. Nous avons pu acquérir des images haute résolution des membres postérieurs inférieurs de souris TDP-43 ΔNLS (rNLS) avec des temps d'acquisition courts. La figure ci-dessous montre le pourcentage de changement du volume musculaire par rapport à la ligne de base à 10 semaines de l'induction de la maladie dans notre modèle de souris « Low Dox ».
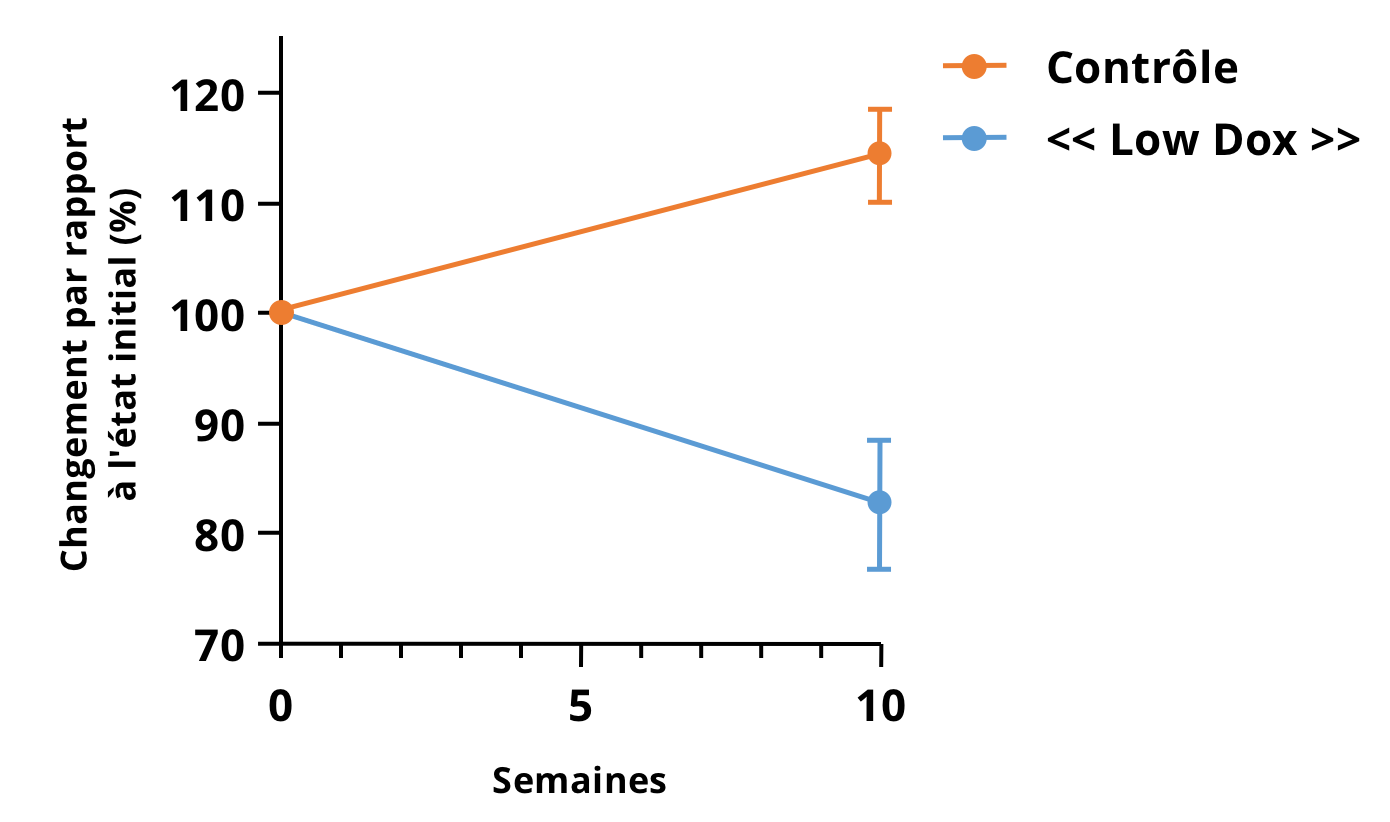
Pourcentage de changement dans les volumes dérivés de l'imagerie micro-CT de divers muscles des membres postérieurs inférieurs dans le modèle de souris hTDP-43ΔNLS « Low Dox ». Notez les différences hautement significatives, en particulier dans les muscles gastrocnémiens à contraction rapide et le StatROI, ce qui rend cette mesure attrayante en tant qu'indicateur de vie pour les études d'efficacité thérapeutique dans ce modèle.
Notez l'atrophie accrue des muscles gastrocnémiens latéraux et médians (à la fois dans le graphique et les cartes) par rapport au soléaire. Rosser et al. (Rosser, 1992) ont montré que la distribution des fibres de type II (à contraction rapide) est la plus élevée dans le gastrocnémien superficiel des rongeurs et la plus faible dans le soléaire.
Ces résultats correspondent bien aux données de Spiller et al. (Spiller, 2016), qui ont rapporté que dans la moelle épinière des souris rNLS8, les « motoneurones lents » ont survécu jusqu'au stade final, tandis que les « motoneurones rapides fatigables » ont été perdus. Ils ont également montré que le dépérissement axonal se produisait d'abord dans les fibres musculaires à contraction rapide, tandis que les fibres à contraction lente restaient innervées. Après la suppression du transgène, les motoneurones résistants ont produit des collatérales pour réinnervé les jonctions neuromusculaires (NMJ) précédemment dénervées.
Electrophysiologie neuromusculaire
L'électrophysiologie (par exemple l'électromyographie [EMG]) est un test standard utilisé chez les patients atteints de SLA pour le diagnostic et le suivi de la maladie. Cette technique peut également être appliquée aux modèles animaux de la SLA pour évaluer l'état fonctionnel neuromusculaire.
Spiller et al. (Spiller, 2016) ont mesuré l'amplitude du potentiel d'action musculaire composé (CMAP) évoqué dans le muscle gastrocnémien après stimulation du nerf sciatique chez des souris non transgéniques (nTg) et rNLS8 qui n'avaient pas reçu de régime à base de doxycycline à haute dose (pour permettre l'expression du transgène) pendant plusieurs semaines. Ce groupe a constaté que les mesures de CMAP diminuaient de manière significative avec l'expression de hTDP-43ΔNLS.
Notre groupe à Biospective a réalisé des études similaires en utilisant notre version « Low Dox » du modèle rNLS8. Conformément aux observations faites dans le modèle « Off Dox », nous avons constaté des réductions hautement significatives sur le plan statistique de l'amplitude de la CMAP dans le modèle « Low Dox ». Ce test peut être réalisé à plusieurs moments et il est bien adapté pour évaluer les effets des agents thérapeutiques expérimentaux dans ce modèle. En outre, l'évaluation de la CMAP complète les mesures de l'atrophie musculaire basées sur l'imagerie micro-CT afin de fournir une image complète des changements neuromusculaires structurels et fonctionnels en fonction de la progression de la maladie et de la modification potentielle de la maladie par le biais d'une intervention thérapeutique.

Traces individuelles montrant une réduction de l'amplitude de la CMAP chez les souris hTDP-43ΔNLS Low Dox par rapport aux souris témoins.
La CMAP a également été utilisée pour évaluer la fonction du système moteur dans les modèles de SOD1 mutants de la SLA chez la souris et le rat. Par exemple, chez les souris SOD1G93A, la CMAP diminue au fil du temps (Mancuso, 2011; Mancuso, 2014). McCampbell et al. (McCampbell, 2018) ont montré que les souris SOD1G93A traitées avec un oligonucléotide antisens (ASO) à l'âge de 5 semaines ont conservé leur CMAP au cours des 12 semaines suivantes, alors que la CMAP des souris témoins a été réduite de plus de la moitié au cours de la même période.
Comment évaluer l'implication des motoneurones spinaux dans les modèles de SLA sur des coupes de tissus?
La visualisation directe, la caractérisation et la quantification des motoneurones spinaux (corps cellulaires et processus) peuvent être réalisées sur des coupes de tissus post-mortem à l'aide de méthodes d'immunohistochimie (IHC) et d'immunofluorescence (IF). L'immunofluorescence multiplexe est particulièrement utile étant donné que plusieurs marqueurs peuvent être évalués sur la même coupe de tissu et que la colocalisation de marqueurs spécifiques peut être évaluée. Une stratégie d'imagerie des motoneurones spinaux sur des coupes de tissus consiste à réaliser un traçage neuronal en injectant des traceurs rétrogrades qui sont soit des molécules fluorescentes (par exemple, la sous-unité B de la toxine cholérique, conjugué Alexa Fluor™ 594 ; CTB-594), soit des vecteurs (par exemple, AAV9) exprimant des gènes rapporteurs (par exemple, GFP, mCherry). Il est également possible d'immunocolorer les antigènes exprimés par les motoneurones spinaux. Les cibles courantes sont la choline acétyltransférase (ChAT), le transporteur vésiculaire d'acétylcholine (VAChT), la métalloprotéinase matricielle-9 (MMP-9) et le canal potassique activé par le calcium de petite conductance de type 3 (SK3).
Une série de mesures quantitatives peut être obtenue à partir de coupes de tissus numérisées. Le nombre et la taille des corps cellulaires et des axones peuvent être mesurés à l'aide de tissus colorés au ChAT ou au VAChT, tandis que les terminaisons peuvent être quantifiées par la coloration du VAChT ou d'autres marqueurs (par exemple SV2). Les marqueurs MMP-9 et SK3 peuvent être utilisés pour évaluer les motoneurones rapides et lents, respectivement. Les mesures morphologiques, telles que la surface du soma et diverses propriétés dendritiques (par exemple, la longueur totale de l'arborescence, la longueur moyenne de l'arborescence, la portée dendritique maximale et la complexité dendritique) (Fogarty, 2020), peuvent également être obtenues à partir d'images de coupes de tissus colorées à l'aide de méthodes avancées de traitement d'images.
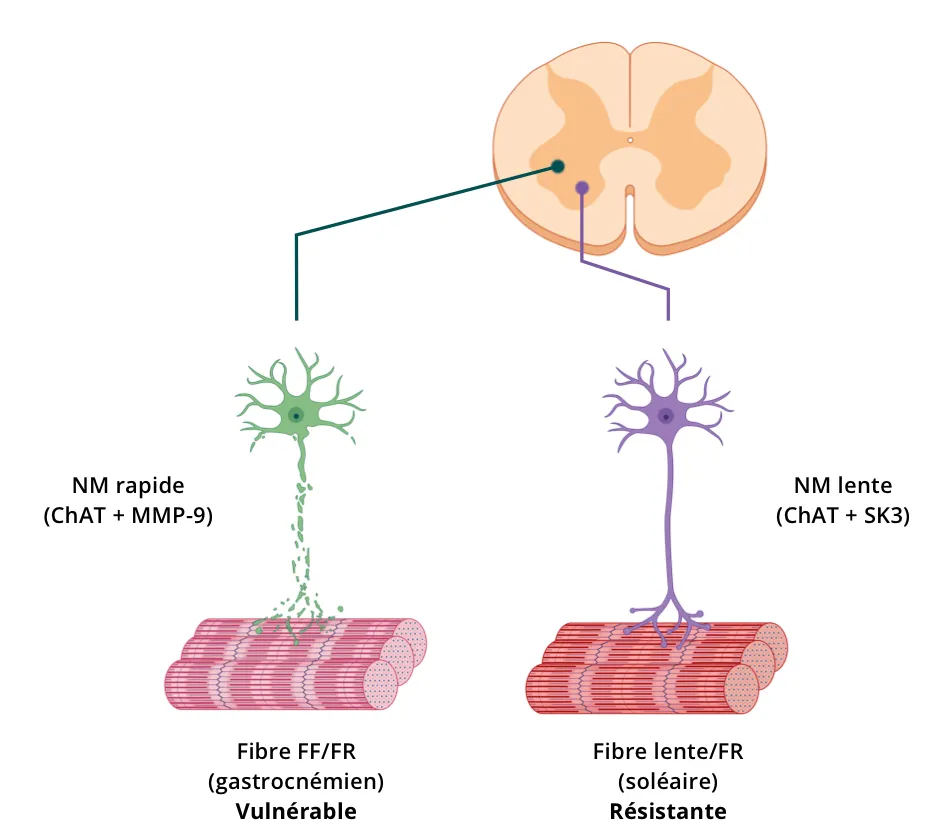
Illustration des stratégies de marquage des motoneurones spinaux dans les modèles animaux (figure reproduite et adaptée de Hur, 2022 sous la Creative Commons Attribution License).
Spiller et al. (Spiller, 2016) ont mesuré le nombre de motoneurones et la taille du soma à l'aide de l'IF VAChT et le nombre de motoneurones rapides via la colocalisation VAChT et MMP-9 dans la moelle épinière de souris hTDP-43ΔNLS. Ce groupe a rapporté que dans la moelle épinière des souris rNLS8, les « motoneurones lents » ont survécu jusqu'au stade final, tandis que les « motoneurones rapides fatigables » ont été perdus. Ils ont également montré que le dépérissement axonal se produisait d'abord dans les fibres musculaires à contraction rapide, tandis que les fibres à contraction lente restaient innervées. Après la suppression du transgène, les motoneurones résistants ont produit des collatérales pour réinnervé les jonctions neuromusculaires (NMJs) précédemment dénervées. La vulnérabilité/résistance sélective des motoneurones peut entraîner une atrophie ou une électrophysiologie différentielle des divers groupes musculaires en fonction de la distribution des types de fibres. Par exemple, le gastrocnémien superficiel est principalement composé de fibres de type II (contraction rapide), tandis que le soléaire est principalement composé de fibres de type I (contraction lente) (Rosser, 1992).
Notre équipe se fera un plaisir de répondre à vos questions sur le modèle de souris TDP-43 ΔNLS ALS ou de vous fournir des informations spécifiques sur les modèles que nous utilisons pour les études d'efficacité thérapeutique.
Découvrez nos modèles ALS
Contenu connexe
Informations actualisées sur la sclérose latérale amyotrophique et les meilleures pratiques liées à l'évaluation des agents thérapeutiques dans les modèles animaux de la SLA.
Autophagie et maladies neurodégénératives
An overview of how cellular autophagy plays a role in brain health and neurodegeneration.
Morphologie de la jonction neuromusculaire (JNM) dans les modèles de SLA
Aperçu de la jonction neuromusculaire (JNM), de son rôle dans la sclérose latérale amyotrophique (SLA) et des outils et méthodes utilisés pour étudier les changements morphologiques dans les JNM.
Modèles de souris SLA pour le développement de médicaments
Un guide pour l'utilisation la plus efficace possible des modèles animaux de recherche sur la sclérose latérale amyotrophique (SLA) pour les essais précliniques de produits thérapeutiques.
Morphologie de la microglie dans la SLA, la maladie d'Alzheimer et la maladie de Parkinson
Une vue d'ensemble de l'analyse morphologique des microglies et des applications à la recherche sur les maladies neurodégénératives et à la découverte et au développement de médicaments.
Analyse de l'atrophie cérébrale dans les modèles murins de neurodégénérescence
Mesures quantitatives automatisées de l'atrophie cérébrale in vivo par IRM (volumes cérébraux régionaux et épaisseur corticale) dans des modèles murins de SLA et de maladie de Parkinson.
Souris TDP-43 ΔNLS (rNLS8) pour le développement de médicaments contre la SLA
Cette ressource fournit des informations sur l'utilisation du modèle de souris transgénique TDP-43 ΔNLS (deltaNLS, hTDP-43ΔNLS, hTDP-43DeltaNLS, dNLS, TDP43 NLS, rNLS8) de la SLA pour des études thérapeutiques précliniques.